Τα πεπτίδια είναι μια κατηγορία ενώσεων που σχηματίζονται από τη σύνδεση πολλαπλών αμινοξέων μέσω πεπτιδίων δεσμών. Είναι πανταχού παρόν σε ζωντανούς οργανισμούς. Μέχρι τώρα, έχουν βρεθεί δεκάδες χιλιάδες πεπτίδια στους ζωντανούς οργανισμούς. Τα πεπτίδια διαδραματίζουν σημαντικό ρόλο στη ρύθμιση των λειτουργικών δραστηριοτήτων διαφόρων συστημάτων, οργάνων, ιστών και κυττάρων και σε δραστηριότητες ζωής και συχνά χρησιμοποιούνται σε λειτουργική ανάλυση, έρευνα αντισωμάτων, ανάπτυξη φαρμάκων και άλλους τομείς. Με την ανάπτυξη της τεχνολογίας βιοτεχνολογίας και πεπτιδίων σύνθεσης, όλο και περισσότερα πεπτιδικά φάρμακα έχουν αναπτυχθεί και εφαρμόζεται στην κλινική.
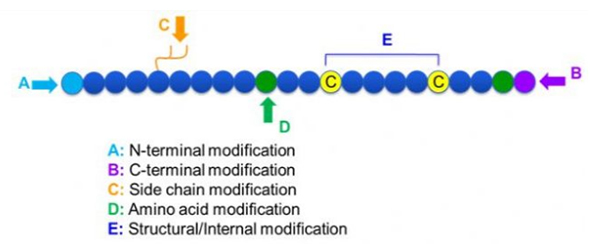
Υπάρχει μια μεγάλη ποικιλία τροποποιήσεων πεπτιδίων, οι οποίες μπορούν απλά να χωριστούν σε τροποποίηση μετά την τροποποίηση και τροποποίηση της διαδικασίας (χρησιμοποιώντας τροποποίηση των προερχόμενων αμινοξέων) και Ν-τερματική τροποποίηση, C-τερματική τροποποίηση, τροποποίηση πλευρικής αλυσίδας, τροποποίηση αμινοξέων, τροποποίηση του σκελετού κ.λπ. ανάλογα με την θέση τροποποίησης (Εικόνα 1). Ως σημαντικό μέσο για την αλλαγή της δομής της κύριας αλυσίδας ή των ομάδων πλευρικής αλυσίδας των πεπτιδίων αλυσίδων, η τροποποίηση πεπτιδίων μπορεί να αλλάξει αποτελεσματικά τις φυσικές και χημικές ιδιότητες των ενώσεων πεπτιδίων, η αύξηση της υδατοδιαλυτότητας, η παρατεταμένη του χρόνου δράσης in vivo, η αλλαγή της βιολογικής κατανομής τους, η εξάλειψη της ανοσογένειας της ανοσογένειας, η μείωση των τοξικών παρενεργειών κλπ. Σε αυτό το χαρτί.
1. Κυκλοποίηση
Τα κυκλικά πεπτίδια έχουν πολλές εφαρμογές στη βιοϊατρική και πολλά φυσικά πεπτίδια με βιολογική δραστηριότητα είναι κυκλικά πεπτίδια. Επειδή τα κυκλικά πεπτίδια τείνουν να είναι πιο άκαμπτα από τα γραμμικά πεπτίδια, είναι εξαιρετικά ανθεκτικά στο πεπτικό σύστημα, μπορούν να επιβιώσουν στην πεπτική οδό και να παρουσιάζουν ισχυρότερη συγγένεια για τους υποδοχείς στόχου. Η κυκλοποίηση είναι ο πιο άμεσος τρόπος συνθέσεως κυκλικών πεπτιδίων, ειδικά για πεπτίδια με μεγάλο δομικό σκελετό. Σύμφωνα με τη λειτουργία κυκλοποίησης, μπορεί να χωριστεί σε τύπο αλυσίδας πλευρικής αλυσίδας, τύπου ακροδεκτών - πλευρικής αλυσίδας, τύπου τερματικού - ακροδέκτη (άκρο προς άκρο).
(1) SideChain-to-Sidechain
Ο πιο συνηθισμένος τύπος κυκλοποίησης πλευρικής αλυσίδας προς πλευρική αλυσίδα είναι η γεφύρωση δισουλφιδίου μεταξύ υπολειμμάτων κυστεΐνης. Αυτή η κυκλοποίηση εισάγεται από ένα ζεύγος υπολειμμάτων κυστεΐνης που αποπληρώνονται και στη συνέχεια οξειδώνεται για να σχηματίσουν δισουλφιδικούς δεσμούς. Η πολυκυκλική σύνθεση μπορεί να επιτευχθεί με επιλεκτική απομάκρυνση ομάδων προστασίας σουλφυδρυλίου. Η κυκλοποίηση μπορεί να γίνει είτε σε έναν διαλύτη μετά την αποκοπή είτε σε ρητίνη προ-διαχωρισμού. Η κυκλοποίηση στις ρητίνες μπορεί να είναι λιγότερο αποτελεσματική από την κυκλοποίηση των διαλύτη επειδή τα πεπτίδια στις ρητίνες δεν σχηματίζουν εύκολα κυκλοποιημένες διαμορφώσεις. Ένας άλλος τύπος κυκλοποίησης πλευρικής αλυσίδας - πλευρικής αλυσίδας είναι ο σχηματισμός μιας δομής αμιδίου μεταξύ ενός ασπαρτικού οξέος ή υπολείμματος γλουταμικού οξέος και του αμινοξέος βάσης, το οποίο απαιτεί η ομάδα προστασίας της πλευρικής αλυσίδας να μπορεί να απομακρυνθεί επιλεκτικά από το πολυπεπτίδιο είτε στη ρητίνη είτε μετά τη διάσπαση. Ο τρίτος τύπος κυκλοποίησης πλευρικής αλυσίδας-πλευρικής αλυσίδας είναι ο σχηματισμός διφαινυλαιθέρων με τυροσίνη ή ρ-υδροξυφαινυλογλυκίνη. Αυτός ο τύπος κυκλοποίησης σε φυσικά προϊόντα βρίσκεται μόνο σε μικροβιακά προϊόντα και τα προϊόντα κυκλοποίησης συχνά έχουν πιθανή φαρμακευτική αξία. Η παρασκευή αυτών των ενώσεων απαιτεί μοναδικές συνθήκες αντίδρασης, επομένως δεν χρησιμοποιούνται συχνά στη σύνθεση συμβατικών πεπτιδίων.

(2) τερματικό σε sidechain
Η κυκλοποίηση της αλυσίδας ακροδεκτών συνήθως περιλαμβάνει το C-τερματικό με την αμινομάδα της πλευρικής αλυσίδας λυσίνης ή ορνιθίνης ή το Ν-τερματικό με την πλευρική αλυσίδα ασπαρτικού οξέος ή γλουταμικού οξέος. Άλλες πολυπεπτιδικές κυκλοποίηση γίνεται με σχηματισμό δεσμών αιθέρα μεταξύ των ακροδεκτών C και σερίνης ή θρεονίνης πλευρικών αλυσίδων.
(3) Τύπος τερματικού ή κεφαλής προς ουρά
Τα πολυπεπτίδια της αλυσίδας μπορούν είτε να κυκλοφορούν σε διαλύτη είτε να σταθεροποιηθούν σε ρητίνη με κύκλους πλευρικής αλυσίδας. Οι χαμηλές συγκεντρώσεις πεπτιδίων θα πρέπει να χρησιμοποιούνται στη συγκέντρωση διαλύτη για να αποφευχθεί η ολιγομερισμός των πεπτιδίων. Η απόδοση ενός πολυπεπτιδίου συνθετικού δακτυλίου κεφαλής προς ουρά εξαρτάται από την αλληλουχία του πολυπεπτιδίου της αλυσίδας. Επομένως, πριν από την παρασκευή κυκλικών πεπτιδίων σε μεγάλη κλίμακα, θα πρέπει πρώτα να δημιουργηθεί μια βιβλιοθήκη πιθανών αλυσιδωτών πεπτιδίων μολύβδου, ακολουθούμενη από κυκλοποίηση για να βρεθεί η ακολουθία με τα καλύτερα αποτελέσματα.
2. Ν-μεθυλίωση
Η Ν-μεθυλίωση εμφανίζεται αρχικά σε φυσικά πεπτίδια και εισάγεται σε πεπτιδική σύνθεση για να αποφευχθεί ο σχηματισμός δεσμών υδρογόνου, καθιστώντας έτσι τα πεπτίδια πιο ανθεκτικά στη βιοαποικοδόμηση και την κάθαρση. Η σύνθεση των πεπτιδίων που χρησιμοποιούν Ν-μεθυλιωμένα παράγωγα αμινοξέων είναι η πιο σημαντική μέθοδος. Επιπλέον, μπορεί να χρησιμοποιηθεί επίσης αντίδραση Mitsunobu του Ν- (2-νιτροβενζολίου σουλφονυλίου χλωριούχου) πολυπεπτιδίου-ερείνης με μεθανόλη. Αυτή η μέθοδος έχει χρησιμοποιηθεί για την παρασκευή κυκλικών πεπτιδικών βιβλιοθηκών που περιέχουν Ν-μεθυλιωμένα αμινοξέα.
3. Φωσφορυλίωση
Η φωσφορυλίωση είναι μία από τις πιο συνηθισμένες μετα-μεταφραστικές τροποποιήσεις στη φύση. Στα ανθρώπινα κύτταρα, πάνω από το 30% των πρωτεϊνών φωσφορυλιώνονται. Η φωσφορυλίωση, ιδιαίτερα η αναστρέψιμη φωσφορυλίωση, παίζει σημαντικό ρόλο στον έλεγχο πολλών κυτταρικών διεργασιών, όπως η μεταγωγή σήματος, η γονιδιακή έκφραση, ο κυτταρικός κύκλος και η ρύθμιση του κυτταροσκελετού και η απόπτωση.
Η φωσφορυλίωση μπορεί να παρατηρηθεί σε μια ποικιλία υπολειμμάτων αμινοξέων, αλλά οι πιο συνηθισμένοι στόχοι φωσφορυλίωσης είναι υπολείμματα σερίνης, θρεονίνης και τυροσίνης. Τα παράγωγα φωσφοτυροσίνης, φωσφορεονίνης και φωσφορίνης μπορούν είτε να εισαχθούν σε πεπτίδια κατά τη διάρκεια της σύνθεσης είτε να σχηματίζονται μετά από πεπτιδική σύνθεση. Η επιλεκτική φωσφορυλίωση μπορεί να επιτευχθεί χρησιμοποιώντας υπολείμματα σερίνης, θρεονίνης και τυροσίνης που απομακρύνουν επιλεκτικά τις προστατευτικές ομάδες. Ορισμένα αντιδραστήρια φωσφορυλίωσης μπορούν επίσης να εισαγάγουν ομάδες φωσφορικών οξέων στο πολυπεπτίδιο με μετα -τροποποίηση. Τα τελευταία χρόνια, η φωσφορυλίωση της λυσίνης έχει επιτευχθεί ειδική για τη θέση χρησιμοποιώντας μια χημικά επιλεκτική αντίδραση staudinger-φωσφορικού (Σχήμα 3).

4. Μυριστοϋλίωση και παλμιτοϋλίωση
Η ακυλίωση του Ν-τερματικού με λιπαρά οξέα επιτρέπει στα πεπτίδια ή τις πρωτεΐνες να δεσμεύονται σε κυτταρικές μεμβράνες. Η μυρριαμοϋλοποιημένη αλληλουχία στο Ν-τερματικό επιτρέπει σε κινάσες πρωτεϊνών οικογενειακής οικογένειας και πρωτεΐνες GAQ ανάστροφης μεταγραφάσης να στοχεύουν να δεσμεύονται με κυτταρικές μεμβράνες. Το μυριστικό οξύ συνδέθηκε με το Ν-τερματικό του πολυπεπτιδίου ρητίνης χρησιμοποιώντας τυπικές αντιδράσεις σύζευξης και το προκύπτον λιποπεπτίδιο θα μπορούσε να διαχωριστεί υπό τυποποιημένες συνθήκες και να καθαριστεί με RP-HPLC.
5. Γλυκοζυλίωση
Τα γλυκοπεπτίδια όπως η βανκομυκίνη και η τεϊγκολανίνη είναι σημαντικά αντιβιοτικά για τη θεραπεία ανθεκτικών σε φάρμακα βακτηριακές λοιμώξεις και άλλα γλυκοπεπτίδια χρησιμοποιούνται συχνά για την τόνωση του ανοσοποιητικού συστήματος. Επιπλέον, δεδομένου ότι πολλά μικροβιακά αντιγόνα είναι γλυκοζυλιωμένα, έχει μεγάλη σημασία να μελετηθούν τα γλυκοπεπτίδια για τη βελτίωση της θεραπευτικής επίδρασης της μόλυνσης. Από την άλλη πλευρά, έχει βρεθεί ότι οι πρωτεΐνες στην κυτταρική μεμβράνη των καρκινικών κυττάρων παρουσιάζουν μη φυσιολογική γλυκοζυλίωση, η οποία καθιστά τα γλυκοπεπτίδια παίζουν σημαντικό ρόλο στην έρευνα για τον καρκίνο και τον όγκο. Τα γλυκοπεπτίδια παρασκευάζονται με μέθοδο FMOC/T-BU. Τα γλυκοζυλιωμένα υπολείμματα, όπως η θρεονίνη και η σερίνη, εισάγονται συχνά σε πολυπεπτίδια από ενεργοποιημένα με FMOCs πενταφθοροφαινόλης για την προστασία γλυκοζυλιωμένων αμινοξέων.
6. Ισοπρένιο
Η ισοπενταδιενυλίωση εμφανίζεται σε υπολείμματα κυστεΐνης στην πλευρική αλυσίδα κοντά στο C-τερματικό. Το ισοπρένιο πρωτεΐνης μπορεί να βελτιώσει τη συγγένεια της κυτταρικής μεμβράνης και να σχηματίσει αλληλεπίδραση πρωτεΐνης-πρωτεΐνης. Οι πρωτεΐνες με ισοπεντραϊτοποιημένες περιλαμβάνουν φωσφατάση τυροσίνης, μικρή GTase, μόρια κοχαπόνεσης, πυρηνικά ελάσματα και πρωτεΐνες κεντρομερούς δέσμευσης. Τα πολυπεπτίδια ισοπρενίου μπορούν να παρασκευαστούν χρησιμοποιώντας ισοπρένιο σε ρητίνες ή εισάγοντας παράγωγα κυστεΐνης.
7. Τροποποίηση πολυαιθυλενογλυκόλης (PEG)
Η τροποποίηση PEG μπορεί να χρησιμοποιηθεί για τη βελτίωση της υδρολυτικής σταθερότητας της πρωτεΐνης, της βιοκατανομής και της διαλυτότητας πεπτιδίου. Η εισαγωγή αλυσίδων PEG στα πεπτίδια μπορεί να βελτιώσει τις φαρμακολογικές τους ιδιότητες και επίσης να αναστέλλει την υδρόλυση των πεπτιδίων από πρωτεολυτικά ένζυμα. Τα πεπτίδια PEG διέρχονται από τη σπειραματική τριχοειδής διατομή πιο εύκολα από τα συνηθισμένα πεπτίδια, μειώνοντας σημαντικά τη νεφρική κάθαρση. Λόγω του εκτεταμένου ενεργού χρόνου ημιζωής των πεπτιδίων PEG in vivo, το φυσιολογικό επίπεδο θεραπείας μπορεί να διατηρηθεί με χαμηλότερες δόσεις και λιγότερο συχνή πεπτιδικά φάρμακα. Ωστόσο, η τροποποίηση PEG έχει επίσης αρνητικές επιπτώσεις. Μεγάλες ποσότητες PEG εμποδίζουν το ένζυμο να υποβαθμίσει το πεπτίδιο και επίσης να μειώσει τη δέσμευση του πεπτιδίου στον υποδοχέα στόχου. Αλλά η χαμηλή συγγένεια των πεπτιδίων PEG αντισταθμίζεται συνήθως από τον μακρύτερο φαρμακοκινητικό τους χρόνο ημιζωής και από το να είναι παρόντες στο σώμα περισσότερο, τα πεπτίδια PEG έχουν μεγαλύτερη πιθανότητα να απορροφηθούν σε ιστούς στόχους. Επομένως, οι προδιαγραφές πολυμερούς PEG θα πρέπει να βελτιστοποιηθούν για βέλτιστα αποτελέσματα. Από την άλλη πλευρά, τα πεπτίδια PEG συσσωρεύονται στο ήπαρ λόγω της μειωμένης νεφρικής κάθαρσης, με αποτέλεσμα το μακρομοριακό σύνδρομο. Ως εκ τούτου, οι τροποποιήσεις PEG πρέπει να σχεδιάζονται πιο προσεκτικά όταν χρησιμοποιούνται πεπτίδια για τη δοκιμή φαρμάκων.

Οι κοινές ομάδες τροποποίησης των τροποποιητών PEG μπορούν να συνοψιστούν κατά προσέγγιση ως εξής: αμινο ( -αμίνη) -ΝΗ2, αμινομεθυλο -CH2 -NH2, υδροξυα -ΟΗ, καρβοξυ -COOH, σουλφυδρυλ ( -θειόλη) -SH, αρσεϊμίδιο -al, ηλεκτρικό ανθρακικό -SC, ηλεκτρικό -succinimide -cmm, suctinimide - n-υδροξυσυκινιμίδη -nhs, ακρυλικό-ch2ch2cooh, αλδεΰδη -Cho (όπως προπιονική-ald, butyrald), ακρυλική βάση (-acrylate-acrl), αζιδο-αζίδη, βιοτινυλο-βιοτίνη, φθορεΐνη, glutaryl -ga, acrylate, alkyne-alkyne p -toluenesulfonate -OTS, succinic -sycinate -ss, κλπ. Παράγωγα PEG με καρβοξυλικά οξέα μπορούν να συνδυαστούν με Ν -τερματικές αμίνες ή πλευρικές αλυσίδες λυσίνης. Το αμινο-ενεργοποιημένο PEG μπορεί να συνδυαστεί με το ασπαρτικό οξύ ή τις πλευρικές αλυσίδες του γλουταμικού οξέος. Το MAL-ενεργοποιημένο PEG μπορεί να συζευχθεί με τη μερκαπτάνη των πλήρως αποπροστατευόμενων πλευρικών αλυσίδων κυστεΐνης [11]. Οι τροποποιητές PEG συνήθως ταξινομούνται ως εξής (Σημείωση: Το MPEG είναι μεθοξυ-ΡΕΕ, CH3O- (CH2CH2O) N-CH2CH2-OH):
(1) Τροποποιητής Peg Straight Chain
MPEG-SC, MPEG-SCM, MPEG-SPA, MPEG-OTS, MPEG-SH, MPEG-ALD, MPEG-Butyrald, MPEG-SS
(2) Διπλουργικός τροποποιητής PEG
HCOO-PEG-COOH, NH2-PEG-NH2, OH-PEG-COOH, OH-PEG-NH2, HCL · NH2-PEG-COOH, MAL-PEG-NHS
(3) Τροποποιητής PEG διακλάδωσης
(MPEG) 2-NHS, (MPEG) 2-ALD, (MPEG) 2-NH2, (MPEG) 2-Mal
8. Βιοτινοποίηση
Η βιοτίνη μπορεί να δεσμεύεται έντονα με άπλαση ή στρεπταβιδίνη και η αντοχή δέσμευσης είναι ακόμη κοντά στον ομοιοπολικό δεσμό. Τα επισημασμένα με βιοτίνη πεπτίδια χρησιμοποιούνται συνήθως σε ανοσοπροσδιορισμό, ιστοκυτταροχημεία και κυτταρομετρία ροής με βάση φθορισμό. Τα ετικέτα αντισώματα αντιβιοτίνης μπορούν επίσης να χρησιμοποιηθούν για τη δέσμευση βιοτινυλιωμένων πεπτιδίων. Οι ετικέτες βιοτίνης συχνά συνδέονται με την πλευρική αλυσίδα λυσίνης ή τον τερματικό Ν. Το 6-αμινοκαπροϊκό οξύ χρησιμοποιείται συχνά ως δεσμός μεταξύ πεπτιδίων και βιοτίνης. Ο δεσμός είναι ευέλικτος στη δέσμευση στο υπόστρωμα και δεσμεύεται καλύτερα με την παρουσία στερεοχημικών παρεμπόδισης.
9. Ετικέτα φθορισμού
Η επισήμανση φθορισμού μπορεί να χρησιμοποιηθεί για την ανίχνευση πολυπεπτιδίων σε ζωντανά κύτταρα και για τη μελέτη των ενζύμων και των μηχανισμών δράσης. Η τρυπτοφάνη (TRP) είναι φθορίζουσα, έτσι ώστε να μπορεί να χρησιμοποιηθεί για εγγενή επισήμανση. Το φάσμα εκπομπών της τρυπτοφάνης εξαρτάται από το περιφερειακό περιβάλλον και μειώνεται με τη μείωση της πολικότητας του διαλύτη, μια ιδιότητα που είναι χρήσιμη για την ανίχνευση της δομής πεπτιδίων και της δέσμευσης υποδοχέα. Ο φθορισμός της τρυπτοφάνης μπορεί να σβήσει με πρωτονιωμένο ασπαρτικό οξύ και γλουταμικό οξύ, το οποίο μπορεί να περιορίσει τη χρήση του. Η ομάδα χλωριούχου δανσυλίου (dansyl) είναι εξαιρετικά φθορίζουσα όταν συνδέεται με μια αμινομάδα και χρησιμοποιείται συχνά ως φθορίζουσα ετικέτα για αμινοξέα ή πρωτεΐνες.
Η μετατροπή ενέργειας συντονισμού φθορισμού (FRET) είναι χρήσιμη για τις μελέτες ενζύμων. Όταν εφαρμόζεται το FRET, το πολυπεπτίδιο του υποστρώματος περιέχει συνήθως μια ομάδα επισήμανσης φθορισμού και μια ομάδα φθορισμού. Οι επισημανόμενες φθορίζουσες ομάδες σβήνονται από τον σβήσιμο μέσω μεταφοράς ενέργειας μη φωτονίων. Όταν το πεπτίδιο διαχωρίζεται από το ενζυμικό ένζυμο, η ομάδα επισήμανσης εκπέμπει φθορισμό.
10. Cage πολυπεπτίδια
Τα πεπτίδια κλουβιών έχουν οπτικά αφαιρούμενες προστατευτικές ομάδες που προστατεύουν το πεπτίδιο από τη δέσμευση στον υποδοχέα. Όταν εκτίθεται σε ακτινοβολία υπεριώδους ακτινοβολίας, το πεπτίδιο ενεργοποιείται, αποκαθιστώντας τη συγγένειά του με τον υποδοχέα. Επειδή αυτή η οπτική ενεργοποίηση μπορεί να ελεγχθεί ανάλογα με το χρόνο, το πλάτος ή τη θέση, τα πεπτίδια κλουβιών μπορούν να χρησιμοποιηθούν για τη μελέτη των αντιδράσεων που εμφανίζονται σε κύτταρα. Οι πιο συχνά χρησιμοποιούμενες προστατευτικές ομάδες για τα πολυπεπτίδια του CAGE είναι 2-νιτροβενζυλικές ομάδες και τα παράγωγά τους, τα οποία μπορούν να εισαχθούν σε σύνθεση πεπτιδίων μέσω προστατευτικών παραγώγων αμινοξέων. Τα παράγωγα αμινοξέων που έχουν αναπτυχθεί είναι η λυσίνη, η κυστεΐνη, η σερίνη και η τυροσίνη. Ωστόσο, τα παράγωγα ασπαρτικού και γλουταμινικού δεν χρησιμοποιούνται συνήθως λόγω της ευαισθησίας τους στην κυκλοποίηση κατά τη διάρκεια της σύνθεσης και της διάστασης πεπτιδίων.
11. Πολυαντογονικό πεπτίδιο (χάρτης)
Τα μικρά πεπτίδια συνήθως δεν είναι ανοσοποιητικά και πρέπει να συνδέονται με πρωτεΐνες φορέα για την παραγωγή αντισωμάτων. Το πολυαντιγονικό πεπτίδιο (MAP) αποτελείται από πολλαπλά πανομοιότυπα πεπτίδια που συνδέονται με πυρήνες λυσίνης, τα οποία μπορούν να εκφράσουν ειδικά ανοσογόνα υψηλής ισχύος και μπορούν να χρησιμοποιηθούν για την παρασκευή ζευγών πρωτεΐνης πεπτιδίου-μεταφορέα. Τα πολυπεπτίδια του MAP μπορούν να συντεθούν με σύνθεση στερεάς φάσης στη ρητίνη MAP. Ωστόσο, η ελλιπής σύζευξη έχει ως αποτέλεσμα τις αλυσίδες που λείπουν ή περικοπή πεπτιδίων σε ορισμένους κλάδους και επομένως δεν παρουσιάζουν τις ιδιότητες του αρχικού πολυπεπτιδίου MAP. Ως εναλλακτική λύση, τα πεπτίδια μπορούν να παρασκευαστούν και να καθαριστούν ξεχωριστά και στη συνέχεια να συνδυαστούν με χαρτογράφηση. Η πεπτιδική αλληλουχία που συνδέεται με τον πεπτιδικό πυρήνα είναι καλά καθορισμένη και εύκολα χαρακτηρίζεται από φασματομετρία μάζας.
Σύναψη
Η τροποποίηση πεπτιδίων είναι ένα σημαντικό μέσο σχεδιασμού πεπτιδίων. Τα χημικά τροποποιημένα πεπτίδια δεν μπορούν μόνο να διατηρήσουν υψηλή βιολογική δραστηριότητα, αλλά και να αποφεύγουν αποτελεσματικά τα μειονεκτήματα της ανοσογονικότητας και της τοξικότητας. Ταυτόχρονα, η χημική τροποποίηση μπορεί να δώσει πεπτίδια με μερικές νέες εξαιρετικές ιδιότητες. Τα τελευταία χρόνια, η μέθοδος της ενεργοποίησης C-H για τη μετα-τροποποίηση των πολυπεπτιδίων έχει αναπτυχθεί ταχέως και έχουν επιτευχθεί πολλά σημαντικά αποτελέσματα.
Χρόνος δημοσίευσης: 2025-07-03